同源四倍体双胚苗材料在其性状表达特征、性状表达频率和性状表达的条件等方面均显现出一定的特异性。在同源四倍体双胚苗材料中,其苗位特征表现出明显的多样性。对于同源四倍体水稻群体内所出现的双胚苗突变材料,按照单株筛选法对其进行2个世代的筛选和纯化,待其主要农艺性状稳定后按照试验设计对其双胚苗的形态特征及其性状稳定性进行了研究。......
2023-11-28
同源四倍体水稻与假稻杂交结籽的胚胎学机制研究(1)
黄群策 向茂成 汤国雄
摘要 利用激光扫描共聚焦显微技术对同源四倍体水稻与假稻杂交结籽的特殊生殖现象进行了研究。研究结果表明,假稻的花粉粒不能在二倍体水稻的柱头上萌发,因而二倍体水稻与假稻的生殖隔离很严格。然而,假稻的花粉粒在同源四倍体水稻的柱头上能萌发,前者的花粉管能在后者的花柱中伸长并能将雄配子送入胚囊内。在同振四倍体水稻与假稻的杂交中受精作用比较特殊,其中包括精细胞与卵细胞的单受精作用。也包括精细胞与欢生极核的单受精作用,还包括正常的双受精作用。总受精率和总成胚率分别为22.89%和20.82%。本试验的结果为解释同源四倍体水稻与假稻杂交结籽的特殊生殖现象提供了有力的胚胎学证据。
关键词 同源四倍体水稻:假稻;激光扫描共聚焦显微技术;胚胎学证据
已经证实,对于一个生物物种而言,其基因资源相当有限,而在整个植物界.其基因资源无穷无尽。通过适当方式打破物种间的生殖隔离,将远缘物种的有益性状引入到普通栽培稻(Oryza sative)中,进而创造出水稻新种质,这对于当前开展的超级稻育种具有重大的意义。假稻(Leerslahexandra)是李氏禾属中的一个物种,具有生长势旺盛、再生能力和宿根能力强、抗多种病虫、耐瘠耐旱等优良性状。同源四倍体水稻(O.sative,2n=4x=48)具有比较弱的有性生殖能力[1,2],它与假稻杂交时表现出一定的可交配性,由此可获得充实度达到正常水平的实粒种子[3]。对于这种在属间有性杂交1中表现出一定的可交配性,不需要进行幼胚拯救就可以获得实粒种子的特殊生殖现象值得进一步研究。本试验以同源四倍体水稻和假稻为杂交亲本,利用激光共聚焦显微技术对其杂交结实的细胞学特征进行了观察和研究,旨在阐明其胚胎学机制,为水稻远缘杂交新技术体系的建立提供细胞学依据。
1 材料和方法
1.1 杂交配组
以同源四倍体水稻紫血稻(4)[试验材料的下标(4)和(2)分别代表同源四倍体水稻和二倍体水稻]和二倍体水稻紫血稻(2)为杂交母本,以假稻为花粉供体进行杂交配组,在授粉前1天对杂交母本的稻穗进行修整,保留第2天将开花的颖花并对其进行人工剪颖去雄。每份材料去雄74个稻穗,在每个稻穗上保留20~30个去雄颖花,去雄后利用羊皮纸袋对其套袋隔离以防串粉。在去雄后的第2天上午向每份母本材料的70个去雄稻穗的颖花内投人新鲜的假稻花粉粒进行授粉除此之外,每份母本材料保留4个去雄稻穗不予授粉以作为对照。
1.2 材料固定
在试验中,在去雄后第2天上午利用新鲜的假稻花粉粒给杂交母本的去雄颖花授粉,随后,按照7个时间段进行材料固定。即在授粉后30min、1.0h、6.0h、24.0h、48.0h、72.0h和94.0h分别从每份母本材料中取10个授粉稻穗(每份材料共计70个授粉稻穗),将其标记后固定在FAA溶液中,在4℃条件下保存备用。
1.3 胚胎学观察
在试验过程中首先将固定在FAA溶液中的颖花取出,剥出其中的子房。在室温条件下分别利用浓度为70%、85%和95%的乙醇对子房进行梯度脱水,在每1梯度乙醇中的浸泡时间为20min。随后,将子房放人无水乙醇中浸泡两次,每次的浸泡时间为2.0h。接着,再转人无水乙醇中过夜。第2天将已经脱水的子房放人由无水乙醇和冬青油(1∶1)组成的混合溶液中过渡1.0h,再用冬青油清洗3次,在前两次清洗时,每次浸泡2.0h,最后一次浸泡的时间为15.0h。所有的试验材料经过冬青油透明处理后可以在冬青油中保存备用。
利用激光扫描共聚焦显微技术对试验材料在授粉后其子房内部的形态特征进行观察鉴定。首先将保存在冬青油中的子房放置在一张凹玻片的中央,加一滴冬青油,盖上盖玻片,再用指甲油封片。随后,在激光扫描共聚焦显微镜(BioRadmRC600 CLSM)下,对每份材料的子房内部结构特征进行观察、分析、比较和鉴定。由此确定试验材料的胚胎学特点,阐明同源四倍体水稻与假稻杂交结籽的胚胎学机制。
2 观察结果及其分析
2.1 花粉粒萌发和花粉管伸长状况
在试验中发现,授粉后假稻的花粉粒不能在二倍体水稻紫血稻(2)的柱头上萌发。在授粉后24.0h的材料中,花粉粒均没有伸出花粉管,并且,在紫血稻(2)的花柱内也没有观察到花粉管。由此可见,二倍体水稻紫血稻(2)不能与假稻进行有性杂交的生殖障碍首先表现在假稻的花粉粒在紫血稻(2)的柱头上不能萌发。此观察结果与我们在进行杂交配组时所得到的结果(即不能获得杂交种子)相吻合[4]。
在本试验中,对已经被去雄隔离48.0h,但没有授予假稻花粉的4个同源四倍体水稻紫血稻(4)的稻穗(共计92朵颖花)进行观察鉴定的结果表明,在各颖花的柱头上没有发现花粉粒,在其花柱内也没有观察到花粉管,由此说明在本试验中的人工剪颖去雄很彻底,套袋隔离也很严格。
在同源四倍体水稻紫血稻(4)与假稻进行杂交的组合中所得到的观察结果则比较复杂。在授粉后30min固定的266枚颖花中,每个颖花的柱头上都沾满着假稻的花粉粒,其中,在21枚颖花(占7.89%)的柱头上发现了正在萌发的花粉粒,其花粉管正伸入柱头内,但尚未伸入到花柱内。在授粉后1.0h固定的306个颖花中,有28个颖花(占9.15%)的花柱内发现了正在伸长的花粉管在授粉后6.0h固定的269枚颖花中,在48个颖花(占17.84%)的花柱内发现了花粉管的痕迹,在19个颖花(占7.06%)的胚囊内的珠孔端发现了花粉管进入胚囊时所留下的痕迹。这些观察结果说明,紫血稻(4)与假稻的生殖隔离不很严格,后者的花粉粒在前者的柱头上可以萌发,花粉管可以在花柱内伸长,直至进入胚囊之内。然而,在紫血稻(4)与假稻的杂交中有2点值得注意。其一是授粉后假稻的花粉粒在紫血稻(4)柱头上的萌发比较缓慢。其二是授粉后花粉管能够进入胚囊的频率比较低。(www.chuimin.cn)
2.2 受精作用和成胚状况
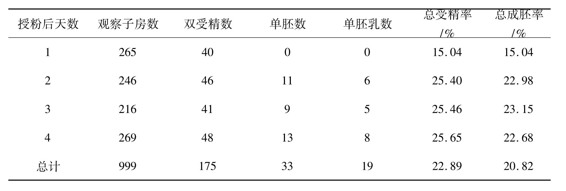
在试验中观察到,假稻的花粉管进入胚囊后,2个精子被释放到卵细胞外的台点端。在授粉后24h固定的一些紫血稻(4)子房中,可以看到卵细胞中出现了雄性核仁,这说明紫血稻(4)的卵细胞与假稻的精细胞发生了两性融合。对授粉后2天、3天和4天的子房分别进行观察鉴定的结果表明,在紫血稻与假稻的杂交中存在着精细胞与卵细胞的单受精现象,精细胞与次生极核的单受精现象和正常的双受精现象。统计结果表明,总受精率高选22.89%,总成胚率也高达20.82%(表1)。这一观察结果与我们在同源四倍体水稻与假稻的杂交试验中所得到的结果相吻合[3]。
表1 紫血稻(4)假稻的受精率
3 讨论
我国超级杂交稻育种是否能取得突破性成就在很大程度上取决于我们是否能通过有效的方法创造出一大批具有实用价值的水稻新种质。已知植物经过长期进化之后,每个物种所具有的基因库相当有限,而在整个植物界却存在着无穷无尽的基因资源有待于我们通过有效的方式来挖掘和利用。自从20世纪60年代以来,许多研究者试图将野生植物中的有益性状引入到水稻中,进而创造出水稻新种质,但由于二倍体水稻具有比较强的有性生殖能力,在二倍体水稻与其他远缘物种的杂交中表现出严格的生殖隔离现象,杂交效率相当低[5-7]。然而,已知同源四倍体水稻具有比较特殊的生殖性状.在以同源四倍体水稻为杂交母本的杂交试验中,远缘物种间具有一定的可交配性[7-8],由此可以获得去雄杂交后充实度达到正常水平的实粒种子。本试验的观察结果表明,假稻的花粉粒在二倍体水稻紫血稻(2)的柱头上不能萌发,两者的生殖隔离发生在受精前,属于配子体隔离从另一方面来看,同源四倍体水稻与假稻的属间杂交则表现出一定的可交配性。在授粉后,假稻的花粉粒在同源四倍体水稻紫血稻(4)的柱头上能萌发,花粉管能伸入花柱内,直至进入胚囊,将两个精细胞释放到卵细胞外的近合点端;在同源四倍体水稻与假稻的杂交中,受精作用比较特殊,存在着正常的双受精。精细胞与卵细胞的单受精和精细胞与次生极核的单受精,总受精率高达22.89%,总成胚率高达20.82%,这些试验结果为同源四倍体水稻与假稻杂交结实的特殊生殖现象提供了有力的胚胎学证据。因此,由于同源四倍体水稻具有比较特殊的生殖特性,它是水稻远缘杂交中的良好亲本和桥梁,由此可望将远缘物种的有益性状转育到水稻中,进而创造出水稻新种质。
References
[1]huang Qunce.Apomous in Angiosperm[M].Fuzhou: Fujian Science Technology Press,2000,3.
[2]Huang Qunce,Sun Jingsan.Bai ShulanStudy on reproductive characters of autotetraploid rice[J].Science Agriculure Sinence,1999.39(2); 14-17.
[3]Huang Qunce.Seeds from autotetraploid rice(Oryza sativa)×heerisahexandra Acta[J].Agrunculture Sinca.2001,27(1):133-135.
[4]HuangQunce,DengQiyun,LiuWenhai.Cytological observation on autotetraploidy rice×Pervisetum alopecurvide.Proceeding of the First Chinese Postdoctoral Symposium[M].Beijing: Science Press,2001,6.
[5]Wu shuxuan,Cai Qigui.Cytological observation onhvbrid F1 from rice[J].Pervisetum alopecurvide Acta Botanica Sinca,1963.11(4):293-301.
[6]Wan Jinju.Sun JingsanCytological observation onhybrid Fl from rice×Pervisetum alopecurvide[J].Acta Botanica Sinca,1981,23(2); 104-109.
[7]Yang Changtao,Xiu Zhongde.Study On obtaining seedings from culture of young embryo in rice distanthybridization[J].Fujian Agriculture Science Technology,1988,13(3):0-3.
[8]Huang Qunce.Effect of Oryza sativa×Pervisetum alopecurvide[J].Chinese Rice Sci,2000,l4(i); 48-59.
【注释】
(1)该文曾在《作物学报》 [2002,28(2):286~288]刊出
有关水稻染色体组多倍化研究的文章

同源四倍体双胚苗材料在其性状表达特征、性状表达频率和性状表达的条件等方面均显现出一定的特异性。在同源四倍体双胚苗材料中,其苗位特征表现出明显的多样性。对于同源四倍体水稻群体内所出现的双胚苗突变材料,按照单株筛选法对其进行2个世代的筛选和纯化,待其主要农艺性状稳定后按照试验设计对其双胚苗的形态特征及其性状稳定性进行了研究。......
2023-11-28

关于多倍体水稻的诱导技术前人已经进行过一些研究[7,8],而关于水稻多倍化的诱导效果与其原始材料的关系的研究目前尚未见到公开的文献报道。对每份材料中所获得的同源四倍体水稻植株的数量进行统计分析,计算多倍化的诱导效果。由此可见,利用常规水稻品种和光温敏核不育水稻品系为诱导材料,其多倍化的诱导频率均很低。利用杂种F1群体为试验材料经过多倍化诱导和筛选后所获得的......
2023-11-28

我国超级稻育种的技术性突破促使稻属遗传改良的水平提升到新的研究层次并给水稻生产带来了革命性的巨变。通过现代生物技术进一步挖掘稻属植物的增产潜力和提高遗传改良水平已经成为水稻育种中的研究热点[7]。其二,在水稻遗传改良的研究领域内研究范围的局限性明显地限制着这一学科在挖掘稻属植物杂种优势效应上发生根本性突破。......
2023-11-28

同源四倍体水稻与假稻杂交结籽的胚胎学机制研究黄群策向茂成汤国雄摘要利用激光扫描共聚焦显微技术对同源四倍体水稻与假稻杂交结籽的特殊生殖现象进行了研究。研究结果表明,假稻的花粉粒不能在二倍体水稻的柱头上萌发,因而二倍体水稻与假稻的生殖隔离很严格。由此确定试验材料的胚胎学特点,阐明同源四倍体水稻与假稻杂交结籽的胚胎学机制。在同源四倍体水稻紫血稻与假稻进行杂交的组合中所得到的观察结果则比较复杂。......
2023-11-28

然而,在千粒重和结实率上,不同染色体组倍性的籼粳亚种间杂种第一代所表现出的差异特别明显。同源四倍体籼粳亚种间杂种第一代在千粒重和结实率上所表现出的特点是挖掘其产量潜力的性状基础。关于同源四倍体籼粳亚种间杂种第一代的产量潜力问题近年来已经引起了一些研究者的关注,但尚未见到公开的文献报道。......
2023-11-28

水稻多倍化及其同源多倍体的潜在价值黄群策代西梅摘要在水稻遗传改良领域我们面临着两大难题需要探索,即如何在促进稻属植物在物种升级的过程中进一步提高其产量潜力和怎样才能固定稻属植物的杂种优势效应。多倍体水稻包括同源多倍体水稻、异源多倍体水稻和同源异源多倍体水稻等多种类型。从同源多倍体水稻的研究现状来看,存在着三大......
2023-11-28

一系法杂交水稻研究的技术策略探讨黄群策摘要通过一系法利用和固定水稻的远缘杂种优势是一项具有极大诱惑力但难度颇大的科研难题。由于多倍体水稻的有性生殖能力明显变弱,在多倍性水平筛选和创造水稻无融合生殖种质有可能成为一系法杂交稻研究的突破口。目前,我国水稻无融合生殖的研究仍然停留在探索性阶段,其首要任务就是要通过采用新的技术路线和策略尽快筛选出具有育种价值的无融合生殖种质。......
2023-11-28

研究结果表明,不同倍性的普通栽培稻与非洲栽培稻之间不存在严格的生殖隔离,通过有性杂交可以获得杂种第一代种子,但其结实率因普通栽培稻的倍性水平而异。由此认为,利用同源四倍体水稻为杂交母本与非洲栽培稻杂交更容易将后者的遗传物质引入到亚洲栽培稻。以同源四倍体水稻为杂交母本,以非洲栽培稻为花粉供体所配制的16个杂交组合均表现出明显的营养生长优势。......
2023-11-28
相关推荐