同源四倍体双胚苗材料在其性状表达特征、性状表达频率和性状表达的条件等方面均显现出一定的特异性。在同源四倍体双胚苗材料中,其苗位特征表现出明显的多样性。对于同源四倍体水稻群体内所出现的双胚苗突变材料,按照单株筛选法对其进行2个世代的筛选和纯化,待其主要农艺性状稳定后按照试验设计对其双胚苗的形态特征及其性状稳定性进行了研究。......
2023-11-28
不同染色体组倍性水稻在谷粒性状上的差异比较(1)
黄雅琴 黄群策 赵帅鹏
摘要 以3份同源四倍体水稻和3份相应的二倍体水稻为材料,对其8个谷粒性状、蛋白质含量和直链淀粉含量进行了差异性比较,通过对谷粒性状间的相关性分析,进一步比较了不同染色体组倍性水稻之间的差异性。结果表明,除了谷粒容重的增减幅度不存在很大差异之外,同源四倍体水稻与相应的二倍体水稻在其他性状的差异性大部分都达到了极显著水平。谷粒体积的变化最大其次是单粒重。染色体组加倍后,谷粒性状间的相关性发生了变化,对单粒重贡献最大的,粒性状有的发生了改变,有的虽然没变,但相关性水平发生了变化。蛋白质含量的增幅(18.52%)比直链淀粉的增幅(2.53%)更明显。
关键词 同源四倍体水稻;谷粒性状;遗异;营养价值
从整个植物界物种的系统发育过程来看,染色体组的多倍化促进了大多数物种不断地由低级生命形式向高级生命形式演化,因此,一些研究者将植物多倍化物种的形成和演化视为物种进化的重要途径之一[1]。当今世界上种植的很多作物,如小麦、燕麦、棉花、烟草及甘蔗等都是多倍体[2]。被子植物中,约70%的种类在进化史中曾经发生过一次或多次多倍化过程[3],且多倍体由于基因组增加,可能隐含更大的改良潜力和更广的适应能力,很多方面具有比二倍体更大的优势,如繁茂性、硕大性、适应力强和生物产量高等优点。稻属植物的染色体组多种多样,亚洲栽培稻均为二倍体(2n=2x=24)。同源四倍体水稻是人工创造的新种质,具有很强的遗传可塑性和很弱的遗传保守性,不但可以促进物种升级和创造出新物种,还有可能在更高层次上利用和固定其强大的杂种优势[4]。目前,同源四倍体水稻与二倍体水稻的差异研究主要集中在生物学特征、生殖发育特性、胚乳蛋白表达、分子标记等方面[5-7],在谷粒性状上的差异还未见报道。
谷粒性状不仅影响千粒重的大小[8],而且还与稻米品质密切相关[9]。稻米的直链淀粉含量与米饭的食味品质有着密切的关系,直链淀粉含量高,米饭硬,黏性小,饭粒干燥而蓬松,色泽较暗;相反,米饭软,黏性大,饭粒光泽度好[10];人类饮食结构的调整对米中蛋白质的含量也提出了新的要求,选育蛋白质含量高的优质米新品种也一直是育种家的目标。因此,对3份不同染色体组倍性的水稻种子的谷粒性状进行研究,旨在研究供试材料在籽粒粒形、谷粒蛋白质含量和直链淀粉含量上的差异性表达特点。
1 材料和方法
1.1 供试材料
试验中所采用的材料是3份同源四倍体水稻[紫粳(4)、圭630(4)、08-04-602]和3份相应的二倍体水稻[紫粳(2)、圭630(2)、08-02-602]。同源四倍体水稻材料分别来源于以相应的二倍体水稻为材料,用秋水仙素对其种芽进行诱导,在诱导后代中筛选到的主要农艺性状稳定的同源四倍体品系[11]。
1.2 试验方法
将每个材料手工去颖壳,数取1 000粒,测定千粒重。随机取饱满谷粒50粒测量其长(×1)、宽(×2)、厚(×3)、单粒重(y),宽、厚均测量谷粒最宽最厚处,精确至0.02mm,重复3次。计算4个2级数据:长宽比(×4)、长厚比(×5)、谷粒体积(×6,以长×宽×厚值近似代替)及谷粒容重(×7,单粒重/谷粒体积)。试验材料的蛋白质含量和直链淀粉含量用美国Foss公司产NIRsystems 5000光栅型近红外分析仪测定。用DPS数据处理软件进行两组平均数student t检验并分析各性状的差异显著性;为了比较不同谷粒性状对单粒重的贡献大小,以单粒重为依变量,其余7个谷粒性状为自变量,进行逐步回归分析,分析谷粒性状间的相关性。
2 结果与分析
2.1 谷粒性状的差异比较
从外观上观察,同源四倍体水稻的籽粒明显比二倍体籽粒大。如图1所示,四倍体的千粒重比二倍体明显增加,紫粳(4)增加了84.19%,圭630(4)增加了63.76%,08-04-602增加了43.56%,其中以紫粳(4)增幅最大。从表1看出,除紫粳的谷粒厚、谷粒容重和圭630的谷粒容重这3项差异不显著,08-602的谷粒容重为5%显著外,其余均为极显著水平。就四倍体的变化趋势而言,除在长厚比上,圭630(4)比圭630(2)减少了8.06%;在谷粒容重上,圭630(4)比圭630(2)减少了1.27%,08-04-602比08-02-602减少了6.33%外,其余的变化趋势均为四倍体较二倍体有所增加,增加率如表1所示。在所观察比较的8个谷粒性状中,谷粒体积的增加幅度最大,平均为76.66%,其次是单粒重,平均为72.66%;谷粒容重的变化最小,平均为41.03%,变幅在1.27%~6.33%。由此可见,同源四倍体水稻比二倍体水稻籽粒更大、更重,同时谷粒容重稳定,极具增产潜力。
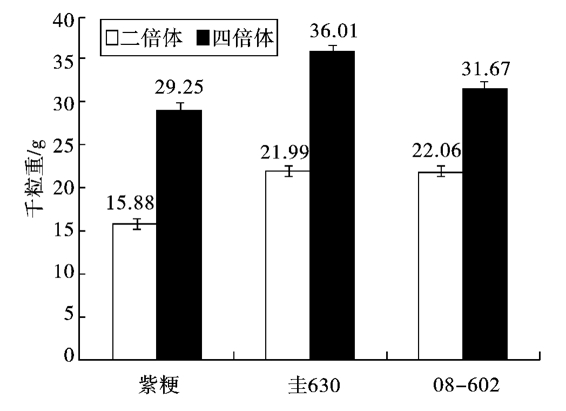
图1 不同倍性水稻的干重差异比较
表1 同材料谷粒性状的差异比较
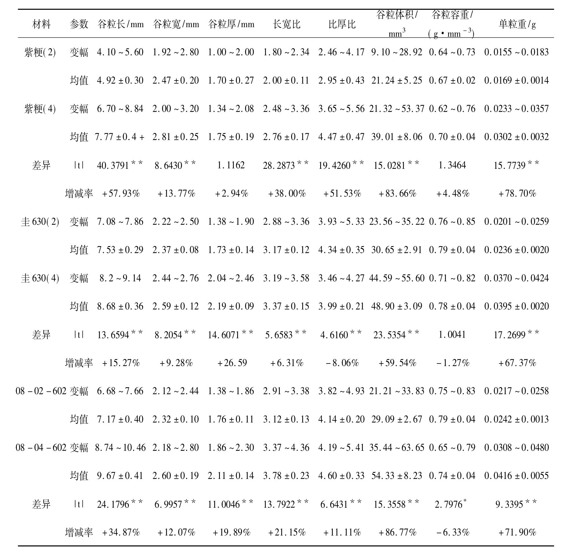
注:*、**分别表示差异达5%或1%显著水平;“+ ”表示四倍体比二倍体有所增加;“-”表示四倍体比二倍体有所减少
2.2 谷粒性状间的相关分析
2.2.1 不同倍性水稻的差异比较
紫粳(2)的逐步回归方程为: y =-0.104 97 +0.102 17×1 +0.102 18×3-0.103 09×4-0.100 15×6 +1.910 738×7(R =0.196 823,SSE =0.100 04)由方程可知,单粒重与谷粒长、谷粒厚、长宽比、谷粒体积、谷粒容重相关,其中与谷粒长、谷粒厚、谷粒容重呈正相关,与长宽比、谷粒体积呈负相关。单粒重与谷粒长、谷粒厚、谷粒容重的偏相关系数分别为0.769 1、0.728 8、0.874 8。由此可见,对单粒重的贡献是谷粒容重>谷粒长>谷粒厚。
紫粳(4)的逐步回归方程为: y =-0.101 66 +0.100 36×1-0.100 72×2-0.100 66×4 +0.100 07×6 +41.141 16×7(R =0193 76**,SSE =0.100 13)由方程可知,单粒重与谷粒长、谷粒宽、长宽比、谷粒体积、谷粒容重相关,其中与谷粒长、谷粒体积、谷粒容重呈正相关,与谷粒宽、长宽比呈负相关。单粒重与谷粒长、谷粒体积、谷粒容重的偏相关系数分别为0.158 023、0.877 6**、0.835 5**,单粒重与谷粒体积的偏相关系数大于单粒重与谷粒容重的偏相关系数,由此可见,对单粒重的贡献是谷粒体积>谷粒容重>谷粒长。由以上分析可以看出,紫粳(2)与紫粳(4)在8个谷粒性状间的相关表现不一。两者表现一致的是:单粒重都与谷粒长、谷粒容重呈正相关,与长宽比呈负相关。相关表现不一致的是:紫粳(2)的单粒重还与谷粒厚呈正相关,与谷粒体积呈负相关,谷粒容重对单粒重的贡献最大,为显著正相关;紫粳(4)的单粒重还与谷粒体积呈正相关,与谷粒厚呈负相关,谷粒体积对单粒重的贡献最大,为极显著正相关。由此可见,紫粳(2)与紫粳(4)差异明显,对单粒重贡献最大的谷粒性状不同。
2.2.2 圭630(2)与圭630(4)的差异比较
圭630(2)的逐步回归方程为: y =-0.014 8 +0.002 3×1 +0.006 2×2 +0.008 7×3-0.001 9×5(R =0.955 7**,SSE = 0.000 8)由方程可知,单粒重与谷粒长、谷粒宽、谷粒厚、长厚比相关,其中与谷粒长、谷粒宽、谷粒厚呈正相关,与长厚比呈负相关。单粒重与谷粒长、谷粒宽、谷粒厚的偏相关系数分别为0.639 7、0.702 0、0.154 81,均未达到显著相关。通过比较偏相关系数可知,对单粒重的贡献是谷粒宽>谷粒长>谷粒厚。
圭630(4)的逐步回归方程为: y =0.101 01 +0.100 16×1 +0.101 39×2-0.100 53×4-0.100 81×5 +35.164 18×7(R =0.196 823,SSE =0.100 05)由方程可知,单粒重与谷粒长、谷粒宽、长宽比、长厚比、谷粒容重相关,其中与谷粒长、谷粒宽、谷粒容重呈正相关,与长宽比、长厚比呈负相关。单粒重与谷粒长、谷粒宽、谷粒容重的偏相关系数分别为0.643 2、0.911 7**、0.843 23。由此可见,对单粒重的贡献是谷粒宽>谷粒容重>谷粒长。
由以上分析可以看出,圭630(2)与圭630(4)在8个谷粒性状间的相关表现有同有异。两者相关表现一致的是:单粒重都与谷粒长、谷粒宽呈正相关,与长厚比呈负相关;均为谷粒宽对单粒重的贡献最大。相关表现不一致的是:圭630(2)的单粒重还与谷粒厚呈正相关,谷粒宽对单粒重的贡献虽然最大,但相关不显著;圭630(4)的单粒重还与谷粒容重呈正相关,与长宽比呈负相关,谷粒宽对单粒重的贡献最大,且相关极显著。由此可见,圭630(2)与圭630(4)差异明显,对单粒重贡献最大的均为谷粒宽,但相关性水平不同。
2.2.3 08-02-602与08-04-602的差异比较
08-02-602的逐步回归方程为: y =-0.038 1 +0.009 7×3 +0.000 7×6 +28.119 98×7(R =0.905 63,SSE =0.000 9)由方程可知,单粒重与谷粒宽、谷粒体积、谷粒容重呈正相关,与它们的偏相关系数分别为0.152 34、0.813 83、0.662 8。由此可见,对单粒重的贡献是谷粒体积>谷粒容重>谷粒宽。08-04-602的逐步回归方程为: y =-0.033 1-0.008 7×4 +0.005 6×5 +0.000 6×6 +63.887 9×7(R =0.989 8**,SSE =0.001 1)由方程可知,单粒重与长宽比、长厚比、谷粒体积、谷粒容重相关,其中与长厚比、谷粒体积、谷粒容重呈正相关,与长宽比呈负相关。单粒重与长厚比、谷粒体积、谷粒容重的偏相关系数分别为0.6838、0.9545**、0.8977**由此可见,对单粒重的贡献是谷粒体积>谷粒容重>长厚比。
由以上分析可以看出,08-02-602与08-04-602在8个谷粒性状间的相关表现有同有异。两者相关表现一致的是:单粒重都与谷粒体积、谷粒容重呈正相关,且均为谷粒体积对单粒重的贡献最大。相关表现不一致的是: 08-02-602的单粒重还与谷粒宽呈正相关,谷粒体积对单粒重的贡献虽然最大,但仅相关显著; 08-04-602的单粒重还与长厚比呈正相关,与长宽比呈负相关,谷粒体积对单粒重的贡献最大,且相关极显著。由此可见,08-02-602与08-04-602差异明显,对单粒重贡献最大的均为谷粒体积,但相关性水平不同。(www.chuimin.cn)
2.3 蛋白质含量和直链淀粉含量的差异比较
由表2可知,3对材料蛋白质含量的变化都达到了极显著水平,而直链淀粉含量的变化却不一致,紫粳达到极显著,圭630差异不显著,08-602达到5%显著。但值得注意的是,并不总是四倍体的含量高于二倍体。紫粳(4)的蛋白质含量比紫粳(2)的蛋白质含量增加11.16%,直链淀粉含量却减少11.43%;圭630(4)的蛋白质含量比圭630(2)的蛋白质含量减少了5.42%,直链淀粉含量却增加了1156%; 08-04-602比08-02-602的蛋白质含量增加了25.87%,直链淀粉含量也增加了3.49%。由此可见,虽然前人的研究认为,染色体组多倍化会导致植物的同化作用增强,组织内含有比较多的蛋白质、碳水化合物等,但此规律在此并不完全适用也出现了四倍体比二倍体有所降低的情况。
表2 蛋白质含量和直链淀粉含量的差异比较(%)
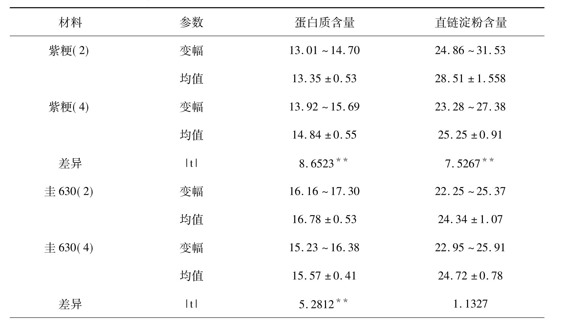
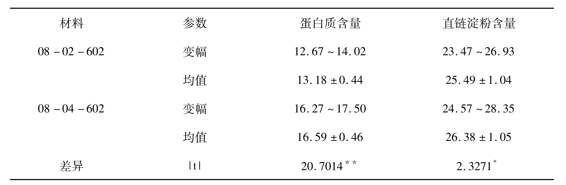
注:*、**分别表示差异达5%或1%显著水平
3 讨论
目前,我国应用于大面积生产的普通二倍体水稻的千粒重多为25 g左右,千粒重的增加能提高产量,同源四倍体水稻以其比二倍体有较大的千粒重等受到了育种家们的关注[13,14],然而,同源四倍体水稻与二倍体水稻相比在谷粒性状上都有哪些差别,这些谷粒性状都与粒重有哪些相关性,如何才能更有效地挖掘同源四倍体水稻的增产潜力,同源四倍体水稻的营养价值又如何,这些问题值得进一步探讨。
本研究结果表明,不同材料除了谷粒容重增减幅度不大外,其他性状大部分都差异极显著,以谷粒体积变化最大,其次是单粒重,这再次证明了同源四倍体水稻的增产潜力。在对谷粒性状间的相关性分析中,分别对3对材料进行了分析比较:对紫粳(2)和紫粳(4)的单粒重贡献最大的分别为谷粒容重和谷粒体积;对圭630(2)和圭630(4)的单粒重贡献最大的都是谷粒宽,但相关水平不同;对08-02-602和08-04-602的单粒重贡献最大的都是谷粒体积,也是相关水平不同。由此可见,要想更有效地挖掘增产潜力,必须针对某一品种进行遗传改良,了解哪些谷粒性状与粒重相关性较大,这样才能为育种工作指明方向,做到有的放矢。
同源四倍体水稻最终要面向市场,其营养价值也是人们所关心的。紫粳(4)与08-04-602的蛋白质含量分别增加了11.16%和25.87%,此增幅与宋文昌等[12]的结果相近。蛋白质含量的增幅与直链淀粉的增幅相比较,蛋白质含量的增幅更加明显,平均为18.52%,而直链淀粉的增幅平均仅为21.53%,较低的直链淀粉含量米饭质地较黏,咀嚼无渣感,有弹性,口感好。因此,染色体组加倍后,在得到较高含量蛋白质的同时又能保证直链淀粉含量不随之大幅度增加从而影响口感,同时还出现紫粳(4)的直链淀粉含量降低11.43%,同源四倍体水稻的这一特性为挖掘其潜在利用价值奠定了基础。
[1]黄群策,秦广雍.禾本科植物染色体组多倍化研究[M].北京:原子能出版社,2008.
[2]Ripolm L,Churchill G A,da Silva JA,et al.Statisticalaspect s of geneticmapping in
autopolyploids[J].Gene,1999,235:31-41.
[3]Masterson J.Stomatal size in fossil plants:evidence forpolyploidy inmajority of angiosperms [J].Science,1994,264:421-424.
[4]黄群策,孙梅元,邓启云.多倍体水稻及其潜在价值[J].杂交水稻,2001,16(1):1-3.
[5]黄群策,孙敬三,白素兰.同源四倍体水稻雌雄配子体多态性的研究[J].杂交水稻,1998,14(1):32-35.
[6]谢慧波,黄群策,李国平,等.不同倍性水稻胚乳蛋白的差异表达研究[J].遗传,2007,29(3):360-364.
[7]栾丽,孔繁伦,何涛,等.用SSR标记检测同源四倍体与二倍体水稻的遗传差异[J].应用与环境生物学报,2004,10(5):556-558.
[8]王余龙,姚友礼,李昙云,等.水稻籽粒有关性状与粒重关系的初步探讨[J].作物学报,1995,21(5):573-578.
[9]杨联松,白一松,张培江,等.谷粒形状与稻米品质相关性研究[J].杂交水稻,2001,16(4):48-50.
[10]陈能,罗玉坤,朱智伟,等.优质食用稻米品质的理化指标与食味相关性研究[J].中国水稻科学,1997,11(2):70-76.
[11]黄群策,代西梅,李玉峰.水稻多倍化的诱导技术研究[J].杂交水稻,2005,20(5):54-56.
[12]宋文昌,张玉华.水稻四倍化及其对农艺性状和营养成分的影响[J].作物学报,1992,18(2):137-144.
[13]汪杏莉,谢慧波,黄群策,等.水稻染色体组加倍对其形态和胚乳蛋白质含量的影响[J].河南农业科学,2008(2):21-24.
[14]黄群策,代西梅,贾宏汝.同源四倍体双胚苗水稻雌配子体的发育特征[J].河南农业科学,2007(4):18-21,51.
【注释】
(1)该文曾在《河南农业科学》 [2009,38(2):57~61]刊出
有关水稻染色体组多倍化研究的文章

同源四倍体双胚苗材料在其性状表达特征、性状表达频率和性状表达的条件等方面均显现出一定的特异性。在同源四倍体双胚苗材料中,其苗位特征表现出明显的多样性。对于同源四倍体水稻群体内所出现的双胚苗突变材料,按照单株筛选法对其进行2个世代的筛选和纯化,待其主要农艺性状稳定后按照试验设计对其双胚苗的形态特征及其性状稳定性进行了研究。......
2023-11-28

稻属植物的进化特点及其潜在价值黄群策王书玉张书艮摘要概括了稻属植物的种群特征及进化特点。关键词稻属植物;种群特征;进化特点;染色体组多倍化;产量潜力在植物物种的自然演化进程中,稻是一种起源比较早、进化程度比较高的古老植物类群。稻属植物是禾本科中非常重要的植物种群,在该属内包含有23个物种,它们广泛分布于全球的热带地区和亚热带地区。......
2023-11-28

我国超级稻育种的技术性突破促使稻属遗传改良的水平提升到新的研究层次并给水稻生产带来了革命性的巨变。通过现代生物技术进一步挖掘稻属植物的增产潜力和提高遗传改良水平已经成为水稻育种中的研究热点[7]。其二,在水稻遗传改良的研究领域内研究范围的局限性明显地限制着这一学科在挖掘稻属植物杂种优势效应上发生根本性突破。......
2023-11-28

其一,氮离子注入后在同源四倍体水稻的后代群体内可以筛选到具有不同特征特性的突变单株,利用离子束生物技术对同源四倍体水稻进行遗传改良的技术路线值得肯定,其实用性已经为同源四倍体水稻新种质的选育找到了新的突破口。......
2023-11-28

关于多倍体水稻的诱导技术前人已经进行过一些研究[7,8],而关于水稻多倍化的诱导效果与其原始材料的关系的研究目前尚未见到公开的文献报道。对每份材料中所获得的同源四倍体水稻植株的数量进行统计分析,计算多倍化的诱导效果。由此可见,利用常规水稻品种和光温敏核不育水稻品系为诱导材料,其多倍化的诱导频率均很低。利用杂种F1群体为试验材料经过多倍化诱导和筛选后所获得的......
2023-11-28
然而,在千粒重和结实率上,不同染色体组倍性的籼粳亚种间杂种第一代所表现出的差异特别明显。同源四倍体籼粳亚种间杂种第一代在千粒重和结实率上所表现出的特点是挖掘其产量潜力的性状基础。关于同源四倍体籼粳亚种间杂种第一代的产量潜力问题近年来已经引起了一些研究者的关注,但尚未见到公开的文献报道。......
2023-11-28

研究结果表明,不同倍性的普通栽培稻与非洲栽培稻之间不存在严格的生殖隔离,通过有性杂交可以获得杂种第一代种子,但其结实率因普通栽培稻的倍性水平而异。由此认为,利用同源四倍体水稻为杂交母本与非洲栽培稻杂交更容易将后者的遗传物质引入到亚洲栽培稻。以同源四倍体水稻为杂交母本,以非洲栽培稻为花粉供体所配制的16个杂交组合均表现出明显的营养生长优势。......
2023-11-28
水稻多倍化及其同源多倍体的潜在价值黄群策代西梅摘要在水稻遗传改良领域我们面临着两大难题需要探索,即如何在促进稻属植物在物种升级的过程中进一步提高其产量潜力和怎样才能固定稻属植物的杂种优势效应。多倍体水稻包括同源多倍体水稻、异源多倍体水稻和同源异源多倍体水稻等多种类型。从同源多倍体水稻的研究现状来看,存在着三大......
2023-11-28
相关推荐