离子注入后同源四倍体核雄性不育水稻的育性表达特性黄群策李玉峰摘要以同源四倍体核不育水稻为研究材料,以氮离子束为诱变源完成了离子注入试验。由此认为,氮离子束注入所导致的育性变异在M3代群体内并没有发生进一步的分离。关于同源四倍体核不育水稻的育性稳定性问题前人已经有过公开的文献报道[3]。利用离子注入技术对同源四倍体核不育水稻进行处理后其雄性不育性的变异特征则是值得研究的问题。......
2023-11-28
同源四倍体光温敏核不育水稻的育性稳定性研究(1)
黄群策 秦广雍 向茂成
摘要 以同源四倍体光温敏核不育水稻为研究材料,对其在不同季节和不同光温条件下的育性稳定性和育性转换时间进行了比较深入的研究。研究结果表明,同源四倍体光温敏核不育水稻具有明显的育性转换特性;无论在夏季的自然条件下还是在人工控制的低温条件下,同源四倍体光温敏核不育水稻比相应的二倍体光温敏核不育水稻表现出更稳定的雄性不育性;前者发生育性转换的起点温度比后者发生育性转换的起点温度要低1.0℃左右;3份同源四倍体光温敏核不育水稻发生育性转换的起点温度均在22.5℃,而二倍体光温敏核不育水稻发生育性转换的起点温度均在23.5℃;在秋季的自然条件下实验中所有的光温敏核不育水稻都能部分结实,结实率的变幅为31.1%~46.9%。
关键词 同源四倍体;光温敏核不育水稻;育性稳定性
在多倍性水平上研究水稻的增产潜力是进一步挖掘和利用水稻杂种优势的重要方面,也是值得深入探索的新领域。我们在研究同源四倍体水稻的生殖特性的过程中已经发现,原始籼稻与原始粳稻在二倍性水平,其杂种第一代的结实率和籽粒充实度很难达到正常水平,而在四倍体水平,其杂种第一代的结实率和籽粒充实度很容易达到正常水平,由此提出了在同源四倍体水平利用原始亚种间强大的杂种优势的新思路[1]。根据两系法杂交稻的育种模式,是否能培育出具有实用价值的光温敏核不育水稻是其技术体系的关键[2]。为了在四倍体水平利用原始籼稻与原始粳稻亚种间强大的杂种优势,我们利用人工诱导法已经将一批二倍体光温敏核不育水稻诱导为同源四倍体光温敏核不育水稻。关于二倍体光温敏核不育水稻的光温生态生理学及其育性稳定性前人已进行过大量的研究[3],而当二倍体光温敏核不育水稻被诱导为同源四倍体光温敏核不育水稻后其育性稳定性和育性转换规律尚未见公开的文献报道。
1 材料和方法
实验中所采用的材料包括3份同源四倍体光温敏核不育水稻品系[培矮64s(4)、99s(4)、7001s(4)]、3份二倍体光温敏核不育水稻品系[培矮64s(2)、99s(2)、7001s(2)]、2份同源四倍体水稻品系[IR36(4)和IR28(4)]以及2份二倍体常规水稻品系(9311和G893)。
鉴定方法包括在自然条件下的鉴定和经过在人工气候箱内进行低温处理后的鉴定。所有的实验材料于2002年3月20日在温室内播种,4月20日移栽至大田。每份材料种植500株,采用常规的田间管理。由于7001s(4)和7001s(2)具有比较强的感光性,因而从4叶期开始对其进行为期4周的遮光处理,以便促进其提早进行幼穗分化,能在夏季高温条件下抽穗开花。从7月20日开始对各份始穗的材料进行定点取稻穗,对其花粉粒育性进行鉴定,每隔5 d取材一次,从每份材料中每次镜检5穗,直至8月23日为止,由此确定各份材料在夏季高温条件下的育性变化状况。同时,对各始穗的材料进行定点套袋隔离,从每份材料中每次套袋隔离5穗,在20 d后对其自交结实率进行统计。从9月20日开始至9月30日为止,每隔2 d取材一次,从每份材料中每次镜检5穗,由此确定各份材料在秋季低温条件下的育性变化状况。同时,对各始穗的材料进行定点套袋隔离,从每份材料中每次套袋隔离5穗,在20 d后对其自交结实率进行统计。
本实验的另一部分就是利用人工气候箱(LRH-250-GS)对每份实验材料在不同温度条件下的育性稳定性和育性转换的临界温度进行鉴定。在人工气候箱内,每日的光照时间为13.5h,光强为5 000 Lx;温度设置为3个日平均温度,即22.0℃、22.5℃和23.5℃,在实验中采用变温处理,相应的温度变化幅度为18.0~24.5℃、18.5~25.0℃和19.5~26.0℃。当实验材料发育到花粉母细胞形成期至减数分裂期这段时期时对其进行人工变温处理。每一批材料在人工气候箱内连续处理6 d后移至田间再让其进一步生长发育。随后,在抽穗期利用1.0%KI-I2溶液对每份材料的花粉粒育性进行鉴定。从每份材料中每次镜检5穗,每穗取10朵颖花混合制片,在光学显微镜下每片取5个视野求平均数,由此统计花粉粒的染色率。同时,对始穗的各份材料进行定点套袋隔离,在20 d后对其自交结实率进行统计。在对实验结果进行统计时,所有已经发生子房膨大但充实度尚未达到正常水平的颖花均被视为结实颖花。
2 结果及其分析
2.1 在夏季自然条件下同源四倍体光温敏核不育水稻雄性不育性的稳定性
在夏季高温自然条件下所有的光温敏核不育水稻,无论是同源四倍体光温敏核不育水稻还是二倍体光温敏核不育水稻,均表现出稳定的雄性不育性,花粉粒的染色率和自交结实率均为0,不育株率和不育度均达到100%。在此条件下,2份同源四倍体水稻品系[IR36(4)和IR28(4)]的花粉粒染色率分别为46.8%和34.9%,自交结实率分别为14.4%和6.7%; 2份二倍体常规水稻品系(9311和G893)的花粉粒染色率分别为88.2%和84.5%,自交结实率分别为86.1%和82.3% (表1)。由此可见,在夏季高温自然条件下实验中所有的光温敏核不育水稻均表现出稳定的雄性不育性。
2.2 在秋季自然条件下同源四倍体光温敏核不育水稻的自交结实性
在秋季低温自然条件下所有的光温敏核不育水稻,无论是同源四倍体光温敏核不育水稻还是二倍体光温敏核不育水稻,均表现出明显的育性转换特性,花药中的一些花粉粒呈现明显的染色反应,花粉可染率的变幅为31.4%~61.3%;稻穗中的部分颖花能结实,结实率的变幅为24.9%~55.2%,其充实度均达到正常水平。在实验中观察到,两种同源四倍体光温敏核不育水稻的育性转换时间明显不同。从9月5日开始,二倍体光温敏核不育水稻先后出现染色的花粉粒,而同源四倍体光温敏核不育水稻在9月14日之后才镜检到染色的花粉粒,这表明同源四倍体光温敏核不育水稻比二倍体光温敏核不育水稻发生育性转换的时间更晚,前者比后者在雄配子或雄配子体的发育上对低温的反应更迟钝。此外,根据对各份材料的自交结实状况的考察结果,同源四倍体光温敏核不育水稻比二倍体光温敏核不育水稻在自交结实率上表现得更差(表2),此问题值得进一步研究。
表1 在夏季自然条件下同源四倍体光温敏核不育水稻雄性不育性的稳定性
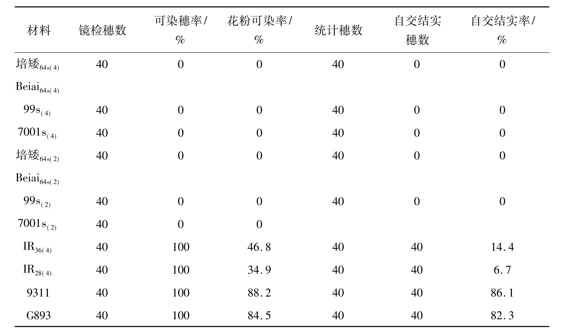
注:考察日期为7月20日至8月23日
表2 在秋季自然条件下同源四倍体光温敏核不育水稻的自交结实性
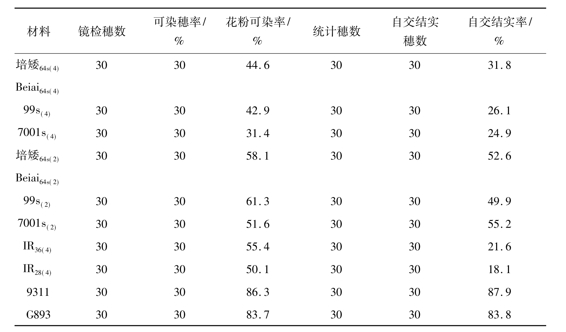
注:考察日期为9月20日至9月30日
2.3在人工控制条件下经过低温处理后同源四倍体光温敏核不育水稻的育性稳定性(www.chuimin.cn)
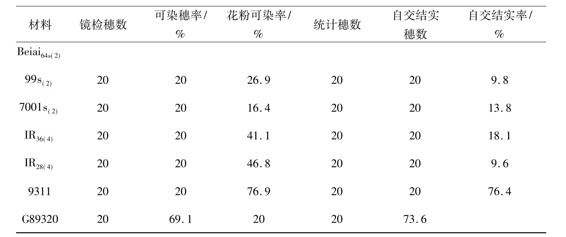
当实验材料发育到花粉母细胞形成期至减数分裂期这段时期内对其进行3种平均温度的人工变温处理后,实验材料在育性表达上的共性很明显,即2份同源四倍体水稻品系[IR36(4)和IR28(4)]和2份二倍体常规水稻品系(9311和G893)对低温处理的反应不是很敏感;同源四倍体光温敏核不育水稻和二倍体光温敏核不育水稻对低温处理的反应却表现出一定的敏感性,但因变温处理中日平均温度的不同而表现出不同的敏感性。在日平均温度为23.5℃的变温处理中,3份同源四倍体光温敏核不育水稻保持着稳定的雄性不育性,其可染穗率、花粉可染率、自交结实穗数和自交结实率均为0,而此条件下3份二倍体光温敏核不育水稻却表现出非常明显的育性转换特性,其花粉粒可染率和自交结实率的变幅分别是16.4%~33.6%和9.8%~13.8%(表3)。在日平均温度为22.5℃的变温处理中,3份同源四倍体光温敏核不育水稻开始表现出明显的育性转换特性,其花粉粒可染率和自交结实率的变幅分别是38.9%~48.1%和31.0%~44.2%,在此条件下3份二倍体光温敏核不育水稻的花粉粒可染率和自交结实率均比较高(表4)。在日平均温度为22.0℃的变温处理中,所有的光温敏核不育水稻,无论是同源四倍体光温敏核不育水稻还是二倍体光温敏核不育水稻都表现出明显的育性转换特性,花粉粒可染率和自交结实率均明显提高,其花粉粒可染率和自交结实率的变幅分别是61.9%~72.8%和51.4%~76.1%。由此可见,在比较低的温度条件下同源四倍体光温敏核不育水稻比相应的二倍体光温敏核不育水稻具有更加稳定的雄性不育性;前者的育性转换起点温度比后者的育性转换起点温度低1.0℃左右; 3份同源四倍体光温敏核不育水稻的育性转换起点温度均在22.5℃,这说明一旦同源四倍体光温敏核不育水稻被应用于生产上进行杂交制种时,如果遇上夏季异常低温,发生育性波动的可能性比较小,雄性不育性比较稳定,其实用价值比较大。然而,尽管在比较低的温度条件下同源四倍体光温敏核不育水稻比相应的二倍体光温敏核不育水稻具有更加稳定的雄性不育性;前者的育性转换起点温度比后者的育性转换起点温度低1.0℃以上,但在自交结实性上前者比后者稍差,这对于同源四倍体光温敏核不育水稻的自交繁殖很不利,对其经济价值有一定的影响,对此问题值得进一步研究。
表3 在人工低温条件下同源四倍体光温敏核不育水稻的自交结实性
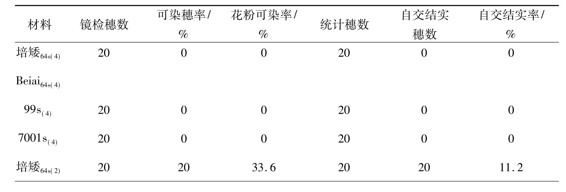
续表3
注:日平均温度为23.5℃,温度变幅为19.5~26.0℃
表4 在人工低温条件下同源四倍体光温敏核不育水稻的自交结实性
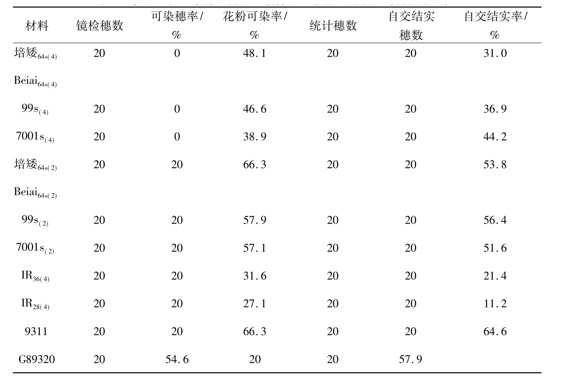
注:日平均温度为22.5℃,温度变幅为18.5~25.0℃
3 讨论
在水稻现有的生产水平上利用新的技术思路和新的研究方法进一步挖掘其增产潜力是水稻育种中值得探索的新领域。过去的研究结果表明,在二倍体水平上普通栽培稻(O.sativa)中原始籼稻和原始粳稻间强大的杂种优势还有待于进一步开发利用。然而,在利用原始籼稻和原始粳稻作为杂交亲本进行杂交稻育种中遇到的最大难题仍然是其杂种第一代的结实率和充实度很难达到正常水平。尽管水稻育种者利用籼粳广亲和性基因可以在一定程度上解决籼粳亚种间杂种第一代的结实率和充实度问题,但由此会使双亲的亲缘关系拉近,遗传距离缩短,因而其杂种优势效应并没有预期的那么强。我们近年来的研究结果表明,将二倍体的原始籼稻和原始粳稻分别诱导为同源四倍体之后,在四倍体水平上原始籼稻和原始粳稻间杂交,其杂种第一代不但具有强大的杂种优势效应,而且其结实率和充实度都会达到正常水平,这预示着在四倍体水平上利用普通栽培稻(O.sativa)中原始籼稻和原始粳稻间强大的杂种优势将是水稻育种中新的研究热点。为了利用原始籼稻和原始粳稻间强大的杂种优势,迫切需要对同源四倍体光温敏核不育水稻的育性稳定性和育性转换特性进行研究。本实验的研究结果表明,同源四倍体光温敏核不育水稻具有明显的育性转换特性;无论在夏季的自然条件下还是在人工控制的低温条件下,同源四倍体光温敏核不育水稻比相应的二倍体光温敏核不育水稻表现出更稳定的雄性不育性;前者发生育性转换的起点温度比后者发生育性转换的起点温度要低1.0℃左右; 3份同源四倍体光温敏核不育水稻发生育性转换的起点温度均在22.5℃,而二倍体光温敏核不育水稻发生育性转换的起点温度均在23.5℃;在秋季的自然条件下实验中所有的光温敏核不育水稻都能部分结实,结实率的变幅为31.1%~46.9%。由此可见,同源四倍体光温敏核不育水稻在利用原始籼稻和原始粳稻间强大的杂种优势中具有一定的实用价值。
[1]黄群策.同源四倍体水稻的潜在价值及其研究策略[J].杂交水稻,2010,9:18-22.
[2]袁隆平.杂交水稻的育种战略设想[J].杂交水稻,1987,2(1):1-3.
[3]周广恰.温敏核不育水稻的光温生态生理学[M].长沙:湖南师范大学出版社,1996.
[4]袁隆平.选育水稻亚种间杂交组合的策略[J].杂交水稻,1996,(2):4-6.
[5]袁隆平.从育种角度展望我国水稻的增产潜力[J].杂交水稻,1996,(4):1-3.
【注释】
(1)该文曾在《作物学报》 [2004,30(2):183~185]刊出
有关水稻染色体组多倍化研究的文章

离子注入后同源四倍体核雄性不育水稻的育性表达特性黄群策李玉峰摘要以同源四倍体核不育水稻为研究材料,以氮离子束为诱变源完成了离子注入试验。由此认为,氮离子束注入所导致的育性变异在M3代群体内并没有发生进一步的分离。关于同源四倍体核不育水稻的育性稳定性问题前人已经有过公开的文献报道[3]。利用离子注入技术对同源四倍体核不育水稻进行处理后其雄性不育性的变异特征则是值得研究的问题。......
2023-11-28
离子注入后同源四倍体多胚苗突变水稻的筛选及其遗传稳定性代西梅黄群策胡秀明摘要以低能氮离子束为诱变源,对同源四倍体水稻品系“IR36-4X”进行离子注入,在其第2代群体内筛选得到了1株具有多胚苗性状特征的突变株。并对该突变株系的苗位、主要农艺性状和多胚苗遗传稳定性进行了跟踪调查。......
2023-11-28

同源四倍体水稻与假稻杂交获得实粒种子黄群策近年来水稻常规育种呈现出停滞不前的状态,这在很大程度上是由于可供育种家利用的有益种质资源不足所致。在本试验中,二倍体水稻与假稻杂交没有获得实粒种子,而同源四倍体水稻与假稻杂交则表现出一定的可交配性,并获得了饱满的实粒种子。......
2023-11-28
第二篇关于同源四倍体水稻的诱导文献综述根据进化论的观点,植物物种的形成和分化要经过漫长的历程,而不同物种在演化过程中所经历的艰难程度都大不相同。其二,在种子露白时进行诱导处理只能使胚芽鞘明显膨大而不能从其后代中筛选到四倍体材料;先行预处理,使胚芽鞘伸长到2.0cm左右时再用秋水仙素对种芽进行诱导处理则能从其后代中筛选到四倍体材料。......
2023-11-28

第五篇关于物种间杂交及同源四倍体水稻的产量潜力文献综述稻属植物是禾本科植物中非常重要的植物种群,它们广泛地分布于全球的热带地区和亚热带地区。由此认为,利用同源四倍体水稻为杂交母本与非洲栽培稻杂交更容易将后者的遗传物质引入亚洲栽培稻。同源四倍体籼粳亚种间杂种第一代在千粒重和结实率上所表现出的特点是挖掘其产量潜力的性状基础。......
2023-11-28

由此可见,利用低能离子注入技术对同源四倍体水稻进行遗传改良的效果值得肯定,这将为解决同源四倍体水稻结实率难题和挖掘其潜在的利用价值寻找到一个新的突破口。根据离子束生物技术的特殊性和实用性,我们以低能氮离子束为诱变源,对同源四倍体水稻进行诱变处理,旨在对其诱变后的生物学效应进行研究,以便筛选出具有实用价值的同源四倍体水稻新种质。......
2023-11-28

在同源四倍体水稻中的子房膨大效应比二倍体水稻中的子房膨大效应更加明显。本文报道了以多个同源四倍体水稻为杂交母本与狼尾草进行杂交,由此获得实粒种子的试验结果。相反,同源四倍体水稻的有性生殖能力比较弱,其遗传保守性比较小而遗传可塑性比较大,利用同源四倍体水稻与狼尾草进行杂交则难度比较小,成功的可能性比较大。......
2023-11-28
其一,氮离子注入后在同源四倍体水稻的后代群体内可以筛选到具有不同特征特性的突变单株,利用离子束生物技术对同源四倍体水稻进行遗传改良的技术路线值得肯定,其实用性已经为同源四倍体水稻新种质的选育找到了新的突破口。......
2023-11-28
相关推荐